
lunes, 25 de marzo de 2019

principales mecanismos
Cladogénesis
La cladogénesis o bifurcación es el mecanismo de especiación más importante. Se produce por aislamiento reproductivo de diferentes poblaciones de una especie debido a las barreras a la hibridación que pueden ser precigóticas o postcigóticas.
Las barreras precigóticas son mecanismos de aislamiento que tienen lugar antes o durante la fecundación, a la que limitan, actúan antes del intercambio gamético. Puede ser por aislamiento ecológico, etológico o mecánico.
Las barreras postcigóticas son todas las que atañen a la viabilidad de los individuos producidos, a través de abortos espontáneos, esterilidad del híbrido, muerte prematura, híbridos débiles y enfermizos, etc.
Hibridación
Hibridación es el cruce reproductivo entre dos especies distintas que pueden producir individuos viables, que pueden ser o no fértiles. Aunque la hibridación en animales puede ser un proceso natural, lo más frecuente es que esté asociado a alteraciones provocadas por la introducción de nuevas especies por el hombre desde que descubrió la ganadería. Es sumamente difícil que esto dé lugar a nuevas especies en los animales.
Entre las plantas la hibridación es un fenómeno extremadamente común. La introgresión es la penetración de genes de una especie en otra por medio de la hibridación. Los híbridos tienden a derivar, en las sucesivas generaciones, hacia los caracteres de uno de los parentales, pero en el proceso puede quedar fijada la transferencia de algunos genes desde la otra especie. No obstante, al igual que en los animales la formación de nuevas especies por hibridación es sumamente rara en las plantas.
Cladogénesis
La cladogénesis o bifurcación es el mecanismo de especiación más importante. Se produce por aislamiento reproductivo de diferentes poblaciones de una especie debido a las barreras a la hibridación que pueden ser precigóticas o postcigóticas.
Las barreras precigóticas son mecanismos de aislamiento que tienen lugar antes o durante la fecundación, a la que limitan, actúan antes del intercambio gamético. Puede ser por aislamiento ecológico, etológico o mecánico.
Las barreras postcigóticas son todas las que atañen a la viabilidad de los individuos producidos, a través de abortos espontáneos, esterilidad del híbrido, muerte prematura, híbridos débiles y enfermizos, etc.
Hibridación
Hibridación es el cruce reproductivo entre dos especies distintas que pueden producir individuos viables, que pueden ser o no fértiles. Aunque la hibridación en animales puede ser un proceso natural, lo más frecuente es que esté asociado a alteraciones provocadas por la introducción de nuevas especies por el hombre desde que descubrió la ganadería. Es sumamente difícil que esto dé lugar a nuevas especies en los animales.
Entre las plantas la hibridación es un fenómeno extremadamente común. La introgresión es la penetración de genes de una especie en otra por medio de la hibridación. Los híbridos tienden a derivar, en las sucesivas generaciones, hacia los caracteres de uno de los parentales, pero en el proceso puede quedar fijada la transferencia de algunos genes desde la otra especie. No obstante, al igual que en los animales la formación de nuevas especies por hibridación es sumamente rara en las plantas.
DEFINICIÓN DEESPECIACIÓN
La noción de especiación no forma parte del diccionario de la Real Academia Española (RAE). El término, de todos modos, es de uso frecuente en el marco de la teoría de la evolución de las especies.
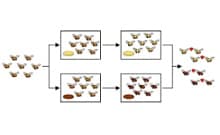
La especiación, en este contexto, es un proceso que hace que la población de una especie propicie la aparición de otra especie diferente. En miles de millones de años, la especiación permitió incrementar de gran forma la diversidad de organismos existentes en el planeta Tierra.
A nivel general, la especiación se vincula a la división de clados o cladogénesis. A través de este proceso, un linaje ancestral termina dividiéndose en varios linajes hermanos, los cuales evolucionan de manera independiente. Por lo tanto, tras la bifurcación inicial, el linaje ancestral queda extinguido y siguen desarrollándose los linajes hermanos en los cuales se concreta la especiación.
Otro mecanismo que deriva en la especiación es la hibridación. En este caso, dos especies diferentes se cruzan y producen nuevos individuos, sean o no fértiles. La hibridación puede desarrollarse de forma natural, aunque también es realizada por el ser humano artificialmente.
El tipo de especiación más habitual es la especiación alopátrica o geográfica. En este caso, una especie se distribuye en un territorio muy amplio, con barreras geográficas como montañas y mares. El acervo genético de la especie, de este modo, queda dividido, estableciéndose diversas poblaciones aisladas que van evolucionando.
La especiación peripátrica, la especiación parapátrica, la especiación simpátrida (también llamada especiación simpátrica) y la especiación cuántica (denominada además especiación saltacional y especiación instantánea) son otros tipos de especiación.
La especiación en las plantas
Por lo que se refiere a la reproducción, las plantas tienen muchas más opciones que los animales. Muchas plantas pueden reproducirse sexualmente, fecundando a otros individuos o a sí mismas, y asexualmente, mediante la creación de clones de sí mismas mediante la reproducción vegetativa, mientras que la mayoría de los animales sólo se reproducen sexualmente.
Igualmente, las plantas tienen más opciones en cuanto a la especiación que los animales. Hay dos tipos de especiación que son particularmente comunes en las plantas:
La especiación mediante hibridación: Por ejemplo, cuando Loren Rieseberg y sus colaboradores reconstruyeron la filogenia de varias especies de girasol, descubrieron que varias especies se habían formado por fecundación entre otras especies. Es frecuente que los descendientes híbridos de tales fecundaciones sean estériles, pero en ocasiones son fértiles y están aislados reproductivamente de sus especies «progenitoras». En este último caso, se forma una nueva especie.




La especie de girasol Helianthus anomalus se formó por la hibridación de otras dos especies de girasol.
Especiación mediante cambios en la ploidía: Desde el punto de vista de la especiación de las plantas, un cambio de ploidía significa, por lo general, la multiplicación del número de cromosomas que tiene la especie por cierto número. Así, una especie que normalmente tiene 18 cromosomas podría producir un linaje que tenga 36 o 54 cromosomas. Los cambios en el número de cromosomas son frecuentes en las plantas y a menudo dan lugar a una especie aislada reproductivamente y diferenciada de la especie parental. Por ejemplo, en la especiación de estas anémonas estuvo implicado un cambio de ploidía.

Aquí vemos dos especies de la flor anémona y sus cromosomas. Los cambios en el número de cromosomas, como ha sucedido en este género, causan la especiación.
Coespeciación
Si la relación entre dos especies es muy cercana, pueden experimentar especiación en paralelo. Se la llama coespeciación, y es especialmente probable que ocurra entre los parásitos y sus hospedadores.
Para ver cómo funciona, imagina una especie de piojo que vive en una especie de ardilla terrestre. Cuando las ardillas se unen para aparearse, el piojo tiene la oportunidad de cambiar de ardilla y, quizás, aparearse con un piojo de otra ardilla. El cambio de ardilla terrestre permite que los genes fluyan en la especie de piojos.

Considera qué le ocurre al piojo si el linaje de ardilla terrestre se divide en los linajes A y B:
El piojo tiene pocas oportunidades de cambiar de ardilla, y el piojo de la ardilla del linaje A no se aparea con el piojo que vive en la ardilla del linaje B.
Este aislamiento «geográfico» de los linajes de piojos puede hacer que lleguen a estar también aislados reproductivamente y, por lo tanto, a ser especies independientes.

Los biólogos evolutivos a menudo pueden decir cuándo los linajes han experimentado coespeciación, ya que la filogenia del parásito «reflejará» la filogenia del hospedador.

El descubrimiento de filogenias paralelas de hospedadores y parásitos es una prueba de la coespeciación.
Este ejemplo está algo idealizado, ya que es muy raro que los científicos encuentren hospedadores y parásitos cuyas filogenias coincidan exactamente. Sin embargo, a veces las filogenias indican que sí que hubo coespeciación, junto con algún cambio de hospedador.
Para ver cómo funciona, imagina una especie de piojo que vive en una especie de ardilla terrestre. Cuando las ardillas se unen para aparearse, el piojo tiene la oportunidad de cambiar de ardilla y, quizás, aparearse con un piojo de otra ardilla. El cambio de ardilla terrestre permite que los genes fluyan en la especie de piojos.
Considera qué le ocurre al piojo si el linaje de ardilla terrestre se divide en los linajes A y B:
El piojo tiene pocas oportunidades de cambiar de ardilla, y el piojo de la ardilla del linaje A no se aparea con el piojo que vive en la ardilla del linaje B.
Este aislamiento «geográfico» de los linajes de piojos puede hacer que lleguen a estar también aislados reproductivamente y, por lo tanto, a ser especies independientes.
Los biólogos evolutivos a menudo pueden decir cuándo los linajes han experimentado coespeciación, ya que la filogenia del parásito «reflejará» la filogenia del hospedador.
El descubrimiento de filogenias paralelas de hospedadores y parásitos es una prueba de la coespeciación.
Este ejemplo está algo idealizado, ya que es muy raro que los científicos encuentren hospedadores y parásitos cuyas filogenias coincidan exactamente. Sin embargo, a veces las filogenias indican que sí que hubo coespeciación, junto con algún cambio de hospedador.
Aislamiento reproductivo
El ambiente puede imponer una barrera externa a la reproducción, como un río o una cordillera, entre dos especies incipientes, pero esa barrera externa por sí sola no las convierte en especies individuales hechas y derechas. La alopatríapuede comenzar el proceso, pero es necesario un desarrollo de barreras internas (es decir, con una base genética) para el flujo génico para que la especiación sea completa. Si no aparecen bareras para el flujo génico, los individuos de las dos partes de la población se aparearán libremente entre ellos si vuelven a entrar en contacto y cualquier diferencia genética que se hubiera producido desaparecerá al volver a mezclarse sus genes. La especiación requiere que las dos especies incipientes sean incapaces de producir descendencia viable juntas o que eviten aparearse con miembros del otro grupo.
Estas son algunas de las barreras para el flujo génico que pueden contribuir a la especiación y que son el resultado de la selección natural, la selección sexual e, incluso, la deriva genética:
El desarrollo de un lugar, un momento o un ritual de apareamiento diferentes: Los cambios con base genética en estos aspectos del apareamiento podrían completar el proceso de aislamiento reproductivo y la especiación. Por ejemplo, los pájaros jardineros (debajo) construyen emparrados elaborados y los decoran con distintos colores para atraer a las hembras. Si dos especies incipientes desarrollaran diferencias en este ritual de apareamiento, podrían quedar aisladas permanentemente y se completaría el proceso de especiación.


Las distintas especies de pájaro jardinero construyen emparrados elaborados y los decoran con colores diferentes para atraer a las hembras. El pájaro jardinero satinado (izquierda) construye un canal entre palos verticales y lo decora con objetos azules brillantes, mientras que el pájaro jardinero de McGregor (derecha) construye una torre de palos alta y la decora con trozos de carbón vegetal. Los cambios evolutivos en los rituales de apareamiento, tales como la construcción del emparrado, pueden contribuir a la especiación.
Falta de «ajuste» entre los órganos sexuales: Es difícil de imaginar para nosotros, pero se trata de un asunto importante para los insectos con genitales de formas variables.
 Estos penes de caballitos del diablo ilustran lo complejos que pueden ser los genitales de los insectos.
Estos penes de caballitos del diablo ilustran lo complejos que pueden ser los genitales de los insectos.Inviabilidad o esterilidad de la descendencia: Todo ese cortejo y apareamiento será en vano si los descendientes de los apareamientos entre los dos grupos no sobreviven o no pueden reproducirse.
En nuestro ejemplo de las moscas-de-la-fruta-en-plátanos-en-putrefacción-en-un-huracán, la alopatría comenzó el proceso de especiación, pero la diferencia de las presiones selectivas en la isla hicieron que la población de la isla se diferenciara genéticamente de la población continental.

El aislamiento geográfico puede provocar un episodio de especiación, pero los cambios genéticos son necesarios para completar el proceso.
¿Cuál pudo ser la causa de esta divergencia? Es posible que las frutas que abundaban en la isla fueran distintas y la población isleña se seleccionara para especializarse en un tipo de fruta determinado y desarrollara una preferencia alimentaria diferente de la de las moscas continentales.

Las diferentes presiones selectivas en las dos islas pueden completar la diferenciación de la nueva especie.
¿Podría esta pequeña diferencia ser una barrera para el flujo génico con las moscas del continente? Sí. Si las moscas encuentran parejas pasando el tiempo en los alimentos preferidos luego, si vuelven al continente, acabarán por no aparearse con las moscas continentales debido a esta preferencia de alimento diferente. El flujo génico disminuiría en gran medida, y una vez que el flujo génico entre las dos especies cesa o disminuye, pueden acumularse diferencias génicas mayores entre las especies.

Pruebas de la especiación
¿La especiación en acción?
En el verano de 1995, al menos 15 iguanas sobrevivieron al huracán Marilyn, sobre una balsa de árboles desarraigados. Viajaron por alta mar durante un mes antes de colonizar la isla caribeña de Anguila. Estos pocos individuos fueron, quizás, los primeros de su especie (Iguana iguana) que llegaron hasta la isla. Si hubo otros intrépidos colonizadores de Iguana iguana en Anguila, se extinguieron antes de que los seres humanos registraran su presencia.
 A los biólogos evolutivos les encantaría saber qué pasa despues: ¿se extinguirán las iguanas colonizadoras?, ¿sobrevivirán y cambiarán sólo un poco?, ¿o terminarán aislandose reproductivamente de las otras Iguana iguana y formarán una nueva especie? Podríamos estar observando los primeros pasos de un suceso de especiación alopátrica, pero en tan poco tiempo no podemos estar seguros.
A los biólogos evolutivos les encantaría saber qué pasa despues: ¿se extinguirán las iguanas colonizadoras?, ¿sobrevivirán y cambiarán sólo un poco?, ¿o terminarán aislandose reproductivamente de las otras Iguana iguana y formarán una nueva especie? Podríamos estar observando los primeros pasos de un suceso de especiación alopátrica, pero en tan poco tiempo no podemos estar seguros.Un modelo plausible: Tenemos varios modelos plausibles sobre cómo sucede la especiación; pero desde luego nos es difícil conseguir un testimonio de algún testigo ocular de un suceso de especiación natural, dado que muchos de estos sucesos ocurrieron en el pasado remoto. Podemos averiguar que han sucedido episodios de especiación y, a menudo, cuándo sucedieron, pero es más difícil averiguar cómo sucedieron. Sin embargo, podemos utilizar nuestros modelos de especiación para hacer predicciones y luego cotejar esas predicciones con nuestras observaciones del mundo natural y los resultados de experimentos. Como ejemplo, vamos a examinar algunas pruebas relevantes del modelo de especiación alopátrica.
Los científicos han encontrado muchas pruebas acordes con que la especiación alopátrica es una forma frecuente de formación de nuevas especies:
Patrones geográficos: Si hay especiación alopátrica, se predeciría que poblaciones de la misma especie en distintas localizaciones geográficas serían genéticamente diferentes. Hay abundantes observaciones que indican que este es un hecho frecuente. Por ejemplo, muchas especies muestran «variedades» regionales que tienen ligeras diferencias genéticas y de apariencia, como es el caso del cárabo californiano del norte y el cárabo californiano del sur. Además, las especies relacionadas son ejemplos convincentes de cómo pueden surgir diferencias genéticas mediante la disminución del flujo génico y la distancia geográfica.

Las subespecies de cárabo californiano que viven en diferentes localizaciones geográficas muestran varias diferencias genéticas y morfológicas: esta observación concuerda con la idea de que las nuevas especies se forman mediante aislamiento geográfico.
Resultados experimentales: En experimentos de laboratorio que incluían aislamiento «geográfico» se han producido los primeros pasos de especiación. Por ejemplo, Diane Dodd tomó moscas de la fruta de una única población y las dividió en poblaciones independientes que vivían en jaulas diferentes para simular aislamiento geográfico. La mitad de las poblaciones vivían de alimentos a base de maltosa y la otra mitad de alimentos a base de almidón. Después de muchas generaciones, se puso a prueba a las moscas para ver con qué otras moscas preferían aparearse. Dodd encontró que se había producido cierto aislamiento reproductivo como consecuencia del aislamiento geográfico y la selección en los distintos ambientes: las «moscas de la maltosa» preferían otras «moscas de la maltosa» y las «moscas del almidón» preferían otras «moscas del almidón». Aunque no podemos estar seguros, es probable que estas diferencias existieran debido a que la selección para la utilización de diferentes fuentes de alimento afectó también a los genes implicados en el comportamiento reproductor. Esta es la clase de resultado que esperaríamos si la especiación alopátrica fuera una forma de especiación típica.

El experimento de la mosca de la fruta de Diane Dodd indica que el aislamiento de poblaciones en ambientes distintos (p. ej., con diferentes fuentes de alimento) puede llevar al comienzo del aislamiento reproductivo. Etos resultados concuerdan con la idea de que el aislamiento geográfico es un paso importante en algunos sucesos de especiación.
Especiación simpátrica
 A diferencia de los tipos anteriores, la especiación simpátrica no requiere una distancia geográfica a gran escala para reducir el flujo génico entre partes de la población.¿Cómo podría reducirse el flujo génico y producirse la especiación en una población que se aparea al azar? La simple explotación de un nuevo nicho puede reducir automáticamente el flujo genético con individuos que explotan el otro nicho. Esto puede suceder ocasionalmente cuando, por ejemplo, insectos herbívoros prueban una nueva planta hospedadora.
A diferencia de los tipos anteriores, la especiación simpátrica no requiere una distancia geográfica a gran escala para reducir el flujo génico entre partes de la población.¿Cómo podría reducirse el flujo génico y producirse la especiación en una población que se aparea al azar? La simple explotación de un nuevo nicho puede reducir automáticamente el flujo genético con individuos que explotan el otro nicho. Esto puede suceder ocasionalmente cuando, por ejemplo, insectos herbívoros prueban una nueva planta hospedadora.Por ejemplo, hace 200 años los antepasados de las moscas de las manzanas ponían sus huevos sólo en las majuelas, pero en la actualidad estas moscas ponen sus huevos en las majuelas (que son originarias de los EEUU) y en las manzanas cultivadas (que fueron introducidas en los EEUU por los inmigrantes y cultivadas). Por lo general las hembras eligen para poner sus huevos el tipo de fruta en el que crecieron y los machos tienden a buscar parejas en el tipo de fruta en el que crecieron. Por lo tanto, las moscas de las majuelas generalmente acaban apareándose con otras moscas de las majuelas y las moscas de las manzanas generalmente acaban apareándose con otras moscas de las manzanas. Esto significa que el flujo génico entre las partes de la población que se aparean en tipos de fruta distintos disminuye. Este cambio de hospedador de las majuelas a las manzanas puede ser el primer paso hacia la especiación simpátrica; en menos de 200 años han evolucionado ciertas diferencias genéticas entre estos dos grupos de moscas.



moscas de las manzanas
manzanas majuelas
El flujo génico ha disminuido entre las moscas que se alimentan de diferentes variedades de alimento incluso aunque ambas viven en la misma región geográfica.
No obstante, los biólogos ponen en duda que este tipo de especiación suceda con mucha frecuencia. En general, la selección para la especialización tendría que ser extremadamente fuerte para hacer que la población se separe, ya que el flujo génico que existe en una población que se aparea al azar tendería a acabar con las diferencias entre las especies incipientes.
Especiación parapátrica
 En la especiación parapátrica no hay ninguna barrera extrínseca concreta para el flujo génico. La población es continua pero, aún así, el apareamiento no es aleatorio. Es más probable que los individuos se apareen con sus vecinos geográficos que con individuos de otra zona del área de distribución de la población. En este tipo de especiación puede haber divergencia debido a una disminución del flujo génico dentro de la población o a presiones selectivas que varían a lo largo del área de distribución de la población.
En la especiación parapátrica no hay ninguna barrera extrínseca concreta para el flujo génico. La población es continua pero, aún así, el apareamiento no es aleatorio. Es más probable que los individuos se apareen con sus vecinos geográficos que con individuos de otra zona del área de distribución de la población. En este tipo de especiación puede haber divergencia debido a una disminución del flujo génico dentro de la población o a presiones selectivas que varían a lo largo del área de distribución de la población. Puede que estemos observando los primeros pasos de especiación parapátrica de la gramínea Anthoxanthum odoratum (a la derecha).
Puede que estemos observando los primeros pasos de especiación parapátrica de la gramínea Anthoxanthum odoratum (a la derecha).Algunas de estas plantas viven cerca de minas donde el suelo se ha contaminado con metales pesados. Las plantas de los alrededores de las minas han estado sometidas a una selección natural de los genotipos tolerantes a los metales pesados, mientras que las plantas vecinas que no viven en suelo contaminado no han estado sometidas a selección para este carácter. Los dos tipos de plantas están lo bastante cerca como para que los individuos tolerantes y los no tolerantes pudieran fecundarse mutuamente, por lo que parece que cumplen el primer requisito de la especiación parapátrica, el de la población continua. Sin embargo, los dos tipos de plantas han desarrollado momentos de floración diferentes. Este cambio podría ser el primer paso en el corte total del flujo génico entre los dos grupos.

Aunque su distribución es continua, los diferentes momentos de floración han hecho que empiece a disminuir el flujo génico entre las plantas tolerantes a los metales y las plantas no tolerantes a los metales.
Especiación peripátrica
 La especiación peripátrica es una versión especial del tipo de especiación alopátrico y sucede cuando una de las poblaciones aisladas tiene muy pocos individuos. He aquí un ejemplo muy hipotético de cómo funciona el tipo de especiación peripátrico. Volviendo a nuestras intrépidas moscas de la fruta que se aventuraban lejos del continente sobre un racimo de plátanos putrefactos, retomemos su historia en el punto en que el racimo es arrastrado hasta una isla:
La especiación peripátrica es una versión especial del tipo de especiación alopátrico y sucede cuando una de las poblaciones aisladas tiene muy pocos individuos. He aquí un ejemplo muy hipotético de cómo funciona el tipo de especiación peripátrico. Volviendo a nuestras intrépidas moscas de la fruta que se aventuraban lejos del continente sobre un racimo de plátanos putrefactos, retomemos su historia en el punto en que el racimo es arrastrado hasta una isla: 1. Un desastre doble: Ahora, las moscas de la fruta de la isla no sólo están aisladas geográficamente de sus parientes del continente, sino que además sólo unas pocas larvas han sobrevivido al vertiginoso viaje y han acabado colonizando la isla.
1. Un desastre doble: Ahora, las moscas de la fruta de la isla no sólo están aisladas geográficamente de sus parientes del continente, sino que además sólo unas pocas larvas han sobrevivido al vertiginoso viaje y han acabado colonizando la isla. 2. Sobreviven genes poco frecuentes: Estos pocos supervivientes tienen, simplemente por casualidad, genes que son poco frecuentes en la población del continente. Da la casualidad de que uno de esos genes poco frecuentes causa una ligera variación en la danza de apareamiento y otro produce una pequeña diferencia en la forma de los genitales masculinos. Este es un ejemplo del efecto fundador.
2. Sobreviven genes poco frecuentes: Estos pocos supervivientes tienen, simplemente por casualidad, genes que son poco frecuentes en la población del continente. Da la casualidad de que uno de esos genes poco frecuentes causa una ligera variación en la danza de apareamiento y otro produce una pequeña diferencia en la forma de los genitales masculinos. Este es un ejemplo del efecto fundador. 3. Las frecuencias génicas derivan:Estas pequeñas diferencias, que son infrecuentes en el continente, derivan hasta quedar fijadas en la pequeña población de la isla en el transcurso de unas pocas generaciones (es decir, la población de la isla al completo acaba teniendo esos genes).
3. Las frecuencias génicas derivan:Estas pequeñas diferencias, que son infrecuentes en el continente, derivan hasta quedar fijadas en la pequeña población de la isla en el transcurso de unas pocas generaciones (es decir, la población de la isla al completo acaba teniendo esos genes). 4. Más cambios: A medida que la población de la isla crece, las características reproductoras de la isla dan lugar a una cascada de cambios causados por la selección sexual. Estos cambios optimizan, o al menos mejoran, la adaptación de los genitales masculinos y femeninos entre sí y la sensibilidad femenina a pequeñas diferencias en el ritual de apareamiento. Las moscas también experimentan selección natural, que favorece a los individuos que mejor se adaptan al clima y la comida de la isla.
4. Más cambios: A medida que la población de la isla crece, las características reproductoras de la isla dan lugar a una cascada de cambios causados por la selección sexual. Estos cambios optimizan, o al menos mejoran, la adaptación de los genitales masculinos y femeninos entre sí y la sensibilidad femenina a pequeñas diferencias en el ritual de apareamiento. Las moscas también experimentan selección natural, que favorece a los individuos que mejor se adaptan al clima y la comida de la isla. 5. Especiación: Después de varias generaciones, las moscas de la isla quedan aisladas reproductivamente de las moscas del continente; se ha producido una especiación peripátrica.
5. Especiación: Después de varias generaciones, las moscas de la isla quedan aisladas reproductivamente de las moscas del continente; se ha producido una especiación peripátrica.En la especiación peripátrica, el pequeño tamaño poblacional hace que la especiación completa sea un resultado más probable del aislamiento geográfico, debido a que la deriva genética actúa más rápido en las poblaciones pequeñas. La deriva genética y, tal vez, unas fuertes presiones selectivas causarían un cambio genético rápido en la pequeña población. Este cambio genético podría conducir a la especiación.
La característica esencial de este tipo es que la deriva genética desempeña un papel en la especiación. Es probable que haya muchos casos en los que una población se divide en dos poblaciones de distinto tamaño y se vuelven especies independientes; sin embargo, es muy difícil para nosotros decir, después de que haya sucedido, cuál fue el papel que desempeñó la deriva genética en la divergencia de las dos poblaciones, por lo que reunir pruevas que apoyen o refuten este tipo de especiación es un reto.
Tipos de especiación
La clave de la especiación es la evolución de diferencias genéticas entre las especies incipientes. Para que un linaje se divida irreversiblemente, las dos especies incipientes deben tener diferencias genéticas que se expresen de alguna forma que haga que no se produzcan apareamientos entre ellas o que, de producirse, sean infructuosos. No hace falta que se trate de enormes diferencias genéticas, un pequeño cambio en el desarrollo cronológico, la localización o los rituales de apareamiento podrían ser suficientes. Pero, eso sí, es necesario que haya alguna diferencia. Este cambio podría producirse por selección natural o por deriva genética.
Es probable que la disminución del flujo génico desempeñe un papel crítico en la especiación. Los tipos de especiación se clasifican normalmente de acuerdo a cuánto contribuye la separación geográfica de las especies incipientes a la disminución del flujo génico. En el siguiente cuadro se comparan algunos de estos tipos de especiación.
La especiación alopátrica: la gran frontera
 La especiación alopátrica es, simplemente, un nombre moderno para la especiación por aislamiento geográfico que vimos anteriormente. En este tipo de especiación algo extrínseco al organismo impide que dos o más grupos se apareen entre sí con regularidad y, finalmente, lleva a la especiación del linaje. El aislamiento puede producirse debido a una gran distancia o a una barrera física, como un desierto o un río, como se muestra debajo.
La especiación alopátrica es, simplemente, un nombre moderno para la especiación por aislamiento geográfico que vimos anteriormente. En este tipo de especiación algo extrínseco al organismo impide que dos o más grupos se apareen entre sí con regularidad y, finalmente, lleva a la especiación del linaje. El aislamiento puede producirse debido a una gran distancia o a una barrera física, como un desierto o un río, como se muestra debajo.
Puede prodicirse especiación alopátrica incluso si la barrera es algo «porosa», es decir, si unos pocos individuos cruzan la barrera y se aparean con miembros del otro grupo. Para que un suceso de especiación se considere «alopátrico», debe haber muy poco flujo génico entre las especies incipientes, pero no tiene que ser completamente inexistente.
Suscribirse a:
Comentarios (Atom)